Transcrição nas E.Coli ( Bactérias)
A transcrição assemelha-se à replicação:
ambos apresentam a mesma direção de síntese, fases de iniciação, alongamento e
terminação. A transcrição difere da replicação pelo fato de não requerer um
iniciador, e apenas um dos lados é usado como molde.
Os trabalhos
e definições das propriedades fundamentais da transcrição tiveram início com o
isolamento e estudo da RNA polimerase. A RNA polimerase depende do
DNA como molde, além de todas as quatros ribunucleosídeos- 5- trifosfato ( ATP, GTP,
UTP, CTP) como precursores das unidades nucleotídicas do RNA,
bem como o íon magnésio. A síntese do RNA tem muito em comum com a síntese do
DNA: A RNA polimerase alonga uma fita de RNA adicionando unidades ribunucleosídeos
na extremidade hidroxila 3’ construindo a fita de RNA na direção 5’-3’.
A iniciação da
síntese do RNA requer várias etapas, geralmente
divididas em duas fases: as fases de ligação e de iniciação. Contudo ela tem
inicio quando
a RNA polimerase se liga a uma sequência especifica do DNA, chamadas
promotores.
A sequência do promotor não é idêntica para todas as bacterianas dessa classe,
dessa forma, as sequências variam de promotor para promotor, todavia, as sequências
em comum são denominadas: sequências
consenso.
Durante
a fase de alongamento
da transcrição, a extremidade crescente da nova fita de RNA faz pareamento de
bases temporariamente com o DNA molde para formar um híbrido RNA-DNA de dupla hélice,
desfaz logo em seguida. Ainda nesta fase à formação das bolhas de transcrição.

Figura
. Transcrição pela RNA polimerase na
E.coli. Para sintetizar uma fita de RNA complementar a uma das duas fitas de
DNA em duplas hélice, o DNA é transitoriamente desenrolado, em um determinado
momento, uma bolha de transcrição é gerada, movendo-se da esquerda para a
direita. O DNA é desenrolado na frente e reenrolado atrás, à medida que o RNA
transcrito.
As duas
fitas complementares do DNA possuem papéis diferentes na transcrição. A fita
que funciona como molde para a síntese do RNA é chamada de fita molde. A fita não de DNA
complementar ao molde, fita-não molde ou codificada que é idêntica na sua sequencia de bases ao RNA transcrito do
gene, com U no lugar de T. O RNA polimerases não possuem uma
atividade exonuclease 3’-5’ revisora, todavia, um erro em uma molécula de RNA é
de menor conseqüência para a célula do que no DNA.
Transcrição nos Eucariotos.
A maquinaria
transcricional no núcleo de uma célula eucariótica é muito mais complexa
do que na bactéria. Os eucariotos possuem três RNA polimerases, designados I, II, III, que são
complexos distintos, mas possuem certas subunidades em comum.
(1) As RNA polimerases
O RNA
Polimerase I (Pol I) é responsável pela síntese de apenas um tipo de RNA, um transcrito de
RNA pré-ribossomal, a principal função da RNA polimerase II (Pol
II) é a sínstese
dos mRNAs e alguns RNAs especializados. O RNA polimerase III ( Pol
III) sintetiza tRNAs,
o rRNA 5S e alguns outros RNAs pequenos especializados.
( Pol
II)
A pol
II é central para a expressão dos genes eucarióticos e tem sido estudada
extensivamente. Ela requer um dispositivo de outras proteínas, chamadas de fatores de
transcrição, ordenadas para formar o complexo de transcrição
ativo. O processo de transcrição pela pol II pode ser descrito em
termos de várias fases- montagem, iniciação, alongamento e terminação. Uma vez
completado o transcrito de RNA, a transcrição é terminada por mecanismos ainda não
reconhecidos. A pol II é desfosforilada e reciclada, pronta para iniciar um
outro transcrito.

Figura . RNA polimerase II
(2) A formação e processamento
do RNA
A formação do RNAm primário ( Pré-síntese)
Uma
molécula
de RNA recém-sintetizada
é chamada de transcrito
primário. O transcrito primário de um mRNA eucariótico usualmente
contém sequencias cercando um gene, embora as sequencias codificadoras do polipeptídio
possa não
ser contínuas. Regiões não-codificadoras que quebram a região
codificadora do transcrito são chamadas de íntrons, e os segmentos codificadores são
chamados de éxons.
Em um processo chamado de emenda, os íntrons são removidos dos transcritos primários
e os éxons unidos para formar uma sequencia continua, especificando um polipeptídio
funcional.

Figura . Pré-síntese do RNA m
Os
mRNAs eucarióticos são também modificados em cada extremidade. Uma estrutura
chamada de capacete
é adicionada à extremidade 5’. A extremidade 3’ é clivada, a alguns
resíduos de adenilato são adicionados para criar uma cauda poli(A)
Complemento: Há
quatro classes de íntrons. As duas primeiras, chamadas de íntrons dos grupos I
e II, diferentes em alguns detalhes de seu mecanismo de emenda. Ambos são autocortáveis, nenhuma enzima está
envolvida neste processo. As funções do capacete 5’ e da cauda poli(A) 3’ são
apenas parcialmente
conhecidas. O capacete 5’ liga-se a uma proteína específica e pode
participar da ligação do mRNA ao ribossomo para iniciar a tradução. A cauda
poli(A) também funciona como um sítio de ligação para uma proteína específica.
É provável
que o capacete 5’, a cauda poli(A) e suas proteínas associadas ajudem a
proteger o mRNA da destruição enzimática. A transcrição dos íntrons
consome recursos celulares e energia, sem retornar nenhum beneficio óbvio ao
organismo; entretanto, os íntrons podem conferir uma vantagem ainda não
considerada pelos cientistas.
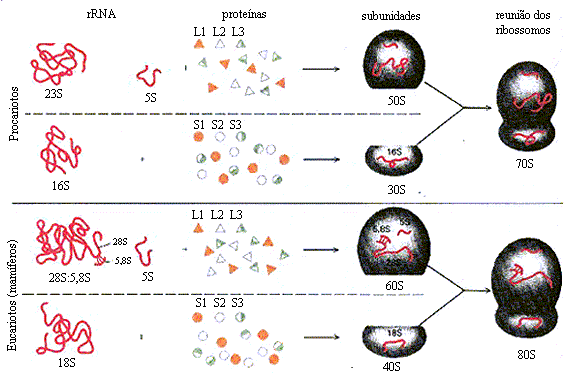
A formação do RNAr e RNAt primário ( Pré-síntese)
O
processamento não apenas limitado ao RNA mensageiro, os RNAs ribossômicos,
tanto bacterianos, quantos eucarióticos, são sintetizados a partir de
precursores denominados: pré-ribossômicos.
Na E.Coli, por exemplo, os RNAs ribossomais
16S, 23S e 5S e alguns transportadores são originados de um único precursor por
múltiplas cópias de gene RNA 30S. Nos eucariotos, um transcrito do RNA
pré-ribossômico 45S é processado no nucléolo para formar os RNAs ribossomais
18S, 28S e 5,8S, característicos dos ribossomos eucarióticos.O
processo de formação então, se dá através da clivagem por ribonucleoproteínas
dos RNAs pré-ribossômicos resultando nos rRNAs tRNAs maduros que vão constituir
o ribossomo e transportadores. A maioria das células possui 40 a 50 tRNAs
distintos, e as células eucarióticas possuem múltiplas cópias de muitos dos
genes de RNA transportador.
Os
transportadores são derivados de precursores de RNA maiores, pela remoção
enzimática das unidades nucleotídicas a partir das extremidades 5’ e 3’ da
molécula madura e retirada do único íntron. O mecanismo de retirada íntron de RNAt
ocorre em duas etapas: na primeira, ocorre a clivagem endonucleasica nas
extremidades 5’-3’. Na segunda etapa ocorre a ligação dos dois exons para a
formação do tRNA maduro. A ligação dos dois exons é feita por um RNA-ligase.
Complemento
Os
precursores de RNA
transportador podem sofrer um outro processamento pós-transcricional.
Este processo é a adição do trinucleotídeo 3’ terminal CCA(3’), ao qual um
aminoácido será ligado durante a síntese de proteínas. Essa adição é realizada
pela enzima tRNA nucleotidiltransferase. Essa definição de nucleotídeos é
independente de um molde de DNA ou RNA.

O tipo final de processamento do tRNA é a modificação de algumas das bases por metilação, deaminação ou redução. Todo o processamento são regulados pela concentração de síntese e degradação dos RNAs. Quando a síntese e a degradação estiverem balanceados, sua concentração permanecerá em equilíbrio estacionário.
Nenhum comentário:
Postar um comentário